


INTRODUCTION
The diversity of insect galls in Neotropical savannas is dependent on both biotic and abiotic factors, although few studies have investigated simultaneously the direct and indirect importance of these factors (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013, Blanche & Westoby Reference BLANCHE and WESTOBY1995). Among the biotic factors that can influence insect gall distribution are the structural parameters of the vegetation (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013). Plant height and abundance, for example, influence positively the amount of resources available for galling insects (as predicted by the resource concentration hypothesis; Root Reference ROOT1973), providing greater area for female oviposition (Cuevas-Reyes et al. Reference CUEVAS-REYES, QUESADA, HANSON, DIRZO and OYAMA2004), and better offspring performance and survival (Santos et al. Reference SANTOS, SILVEIRA and FERNANDES2008). Thus, both plant height and abundance can increase insect gall richness because they maximize the supply of resources and improve conditions for gall-inducing insects (Gonçalves-Alvim & Fernandes Reference GONÇALVES-ALVIM and FERNANDES2001).
Soil fertility is a limiting factor for plant growth in Neotropical savannas (Viana et al. Reference VIANA, RODRIGUES, DAWSON and OLIVEIRA2011, Vourlitis et al. Reference VOURLITIS, LOBO, LAWRENCE, HOLT, ZAPPIA, PINTO and NOGUEIRA2014), affecting plant–herbivore interactions (Marques et al. Reference MARQUES, PRICE and COBB2000, Neves et al. Reference NEVES, ARAÚJO, ESPÍRITO-SANTO, FAGUNDES, FERNANDES, SANCHEZ-AZOFEIFA and QUESADA2010, Zava & Cianciaruso Reference ZAVA and CIANCIARUSO2014). Plant species tend to have lower rates of growth and higher concentrations of defence compounds under poor soil conditions (Neves et al. Reference NEVES, ARAÚJO, ESPÍRITO-SANTO, FAGUNDES, FERNANDES, SANCHEZ-AZOFEIFA and QUESADA2010), which positively influences the performance and survival of galling insects (Fernandes & Price Reference FERNANDES and PRICE1988, Price et al. 1988). A probable explanation for this is that insect galls can sequester the plant secondary metabolites as a mechanism to protect them against natural enemies (Cuevas-Reyes et al. Reference CUEVAS-REYES, QUESADA, HANSON, DIRZO and OYAMA2004, Reference CUEVAS-REYES, OLIVEIRA-KER, FERNANDES and BUSTAMANTE2011). In this context, gall-inducing insects are expected to accumulate more species in environments with lower soil nutrient availability (Blanche & Westoby Reference BLANCHE and WESTOBY1995).
Many previous studies have shown a positive effect of vegetation structure (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013, Cuevas-Reyes et al. Reference CUEVAS-REYES, QUESADA, HANSON, DIRZO and OYAMA2004, Gonçalves-Alvim & Fernandes Reference GONÇALVES-ALVIM and FERNANDES2001) and a negative effect of soil fertility (Cuevas-Reyes et al. Reference CUEVAS-REYES, SIEBE, MARTÍNEZ-RAMOS and OYAMA2003, Gonçalves-Alvim & Fernandes Reference GONÇALVES-ALVIM and FERNANDES2001, Veldtman & McGeoch Reference VELDTMAN and MCGEOCH2003, Wright & Samways Reference WRIGHT and SAMWAYS1996) on the diversity of tropical galling insects. However, few of these studies have considered that the effect of these factors on insect gall diversity may be dependent on plant species richness (Blanche & Westoby Reference BLANCHE and WESTOBY1995). Plant species richness in Neotropical savannas is usually related to plant density and soil fertility (Felfili et al. Reference FELFILI, SILVA-JÚNIOR, SEVILHA, FAGG, WALTER, NOGUEIRA and REZENDE2004, Furley Reference FURLEY1999, Silva et al. Reference SILVA, BATALHA and CIANCIARUSO2013), since these factors favour the accumulation of plant species with more distinct features (Ribeiro & Tabarelli Reference RIBEIRO and TABARELLI2002). Because plant species richness directly affects galling insect richness (review in Araújo Reference ARAÚJO2013), it is expected that part of the effect of vegetation structure and soil fertility on galling insect richness will be dependent on the indirect effects of these variables through plant species richness.
The present study investigates the effects of vegetation structure, soil fertility and plant species richness on insect gall richness in a savanna in Brazil. The initial aim is to test the hypothesis that insect gall richness is higher in vegetation with more complex structure. Abundance and height of plants were used as measures of vegetation structure, because these measures indicate the quantity of available resource (biomass) for galling insects (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013, Cuevas-Reyes et al. Reference CUEVAS-REYES, QUESADA, HANSON, DIRZO and OYAMA2004). The second hypothesis tested is that insect gall richness is higher in environments with lower soil fertility. Soil fertility was measured as the percentage of organic matter and soil macronutrients (N, P, Ca and Mg), which influence plant growth and defence (Cuevas-Reyes et al. Reference CUEVAS-REYES, SIEBE, MARTÍNEZ-RAMOS and OYAMA2003, Reference CUEVAS-REYES, OLIVEIRA-KER, FERNANDES and BUSTAMANTE2011; Zava & Cianciaruso Reference ZAVA and CIANCIARUSO2014). A further objective is to evaluate the extent to which the effects of vegetation structure and soil characteristics on insect gall richness are mediated by indirect effects of these variables via increase in plant species richness.
METHODS
Study area
The study was performed in the Parque Nacional das Emas (17°49′–18°28′S, 52°39′–53°10′W). The Parque Nacional das Emas (PNE) is located in the south-western portion of the state of Goiás, Brazil, with the main entrance located in the city of Mineiros. The climate of the region is classified as Aw of Köppen (Alvares et al. Reference ALVARES, STAPE, SENTELHAS, DE MORAES and SPAROVEK2013), being humid tropical with well-defined dry (April–September) and rainy (October–March) seasons. The majority of the 132941 ha of the park is flat tableland at 820–888 m asl, whereas the remaining area consists of hilly terrain at 720–820 m asl (Ramos-Neto & Pivello Reference RAMOS-NETO and PIVELLO2000). The PNE is located in phytogeographic domain of cerrado, which is the largest savanna of South America and one of the richest savannas in the world (Klink & Machado Reference KLINK and MACHADO2005). The PNE exhibits almost all types of cerrado vegetation, but mainly forest formations (e.g. semideciduous and gallery forests), and savanna formations (e.g. dense and open savannas), with the latter occupying 78.5% of the area of park (Ramos-Neto & Pivello Reference RAMOS-NETO and PIVELLO2000).
Sampling
Sampling was performed in the beginning of the rainy season (November 2013), which is a more favourable period for galling insect occurrence because of the appearance of new host organs such as shoots and leaves (Cuevas-Reyes et al. Reference CUEVAS-REYES, QUESADA and OYAMA2006). Insect galls were collected from 49 10 × 10-m plots distributed in a gradient of open and dense savanna (39 plots in open savanna and 10 plots in dense savanna) in PNE. Woody plants with a diameter at ground level ≥5 cm were identified in the field and had their leaves inspected for insect galls. All insect galls encountered were recorded, photographed, collected and placed individually in labelled plastic bags for transportation to the laboratory. Insect galls were classified into morphotypes using the host plant species and external morphology (organ of occurrence, form, colour, pubescence and size) (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013). Gall morphotypes were used as a surrogate for species of gall-inducing insects because there is a consensus in the literature about host-specificity and morphological-fidelity of insect galls (review in Carneiro et al. Reference CARNEIRO, BRANCO, BRAGA, ALMADA, COSTA, MAIA and FERNANDES2009).
Plant species richness, vegetation structure variables and soil characteristics were described for each sampled plot (Appendix 1). Plant species richness was considered as the number of woody plant species in each plot, and vegetation structure was described using abundance (number of individual plants) and height (mean height of individual plants) for each plot. To further describe each plot, the percentage of organic matter (OM), nitrogen (N), phosphorus (P), calcium (Ca) and magnesium (Mg) were measured. These soil characteristics were determined for five 500-g soil samples taken at 0–10 cm depth in each plot. The five samples for each plot were combined to produce a composite representative for each plot. Soil data were originally collected in the 49 plots by Zava & Cianciaruso (Reference ZAVA and CIANCIARUSO2014), who provide additional information on soil characteristics of the PNE.
Data analyses
A path analysis (Grace Reference GRACE2006) was used to determine the effects of vegetation structure, soil characteristics and plant species richness on insect gall richness. Prior to the construction of path model, multicollinearity between explanatory variables was tested using Pearson correlation (Appendix 2). Aiming to avoid inflating the model with too many explanatory variables, a principal components analysis (PCA) was used to transform the soil variables (N, P, Ca and Mg) into orthogonal components (uncorrelated) that could be used as explanatory variables in the analysis. The first axis of the PCA, which explained over 80% of the variation in the soil characteristics, is here on referred to as soil macronutrients.
For the path analysis the variables of vegetation structure (plant abundance and plant height) and soil characteristics (soil macronutrients and soil organic matter) were included as exogenous explanatory variables. In turn, plant species richness was included in the path model as both an exogenous and an endogenous variable, because it is expected to be influenced by vegetation structure and soil characteristics and to influence insect gall richness. The path model was constructed and analysed using the free version of AMOS 5.0 (Arbuckle Reference ARBUCKLE2003).
Model fit was assessed using chi-squared tests and by examining the Tucker–Lewis Fit Index (TLI), the Comparative Fit Index (CFI), and the Root Mean Square Error of Approximation (RMSEA). These measures are known as confirmatory factor analyses (CFA), which is a special form of factor analysis used to test whether data fit a hypothesized measurement model (Schmitt Reference SCHMITT2011). The approach introduced by MacCallum et al. (Reference MACCALLUM, BROWNE and SUGAWARA1996) was used to verify whether the path model had adequate statistical power. Statistical power was calculated using RMSEA through the R software (Preacher et al. Reference PREACHER, CAI, MACCALLUM, Little, Bovaird and Card2007). According to Schmitt (Reference SCHMITT2011), each goodness-of-fit measure has an adequate value as follows: χ² (close to 0.0), TLI (> 1.0), CFI (> 0.95), RMSEA (< 0.06) and statistical power (close to 1). Normality was checked using the Mardia test, and outliers were inspected using Mahalanobis distance.
RESULTS
Altogether a total of 985 woody plants were sampled comprising 71 species and 25 families, of which 186 individuals, 55 species and 24 families hosted insect galls. A total of 97 insect gall morphotypes was recorded, with Cecidomyiidae (Diptera) being the most common insect taxon having induced 38.1% of the gall morphotypes, whereas Lepidoptera and Hemiptera induced 5.2% and 2.0%, respectively. Gall-inducing insects could not be found for 54.6% of the gall morphotypes. The plant families with the greatest richness of insect galls were Myrtaceae, with 16 morphotypes, and Fabaceae, with 14 morphotypes. Insect galls were more frequent on leaf lamina with 71 morphotypes, followed by midvein (21) and petiole (4). Fifty-nine gall morphotypes (60.8%) were restricted to dense savanna and 56 (57.7%) to open savanna and only 20 morphotypes (20.6%) occurred in both habitats. The mean (± SD) richness of insect galls in each plot was 9.22 ± 5.24 morphotypes.
The path model of insect gall richness explained 73% of the variation in the total number of gall-inducing species (Figure 1). This path model showed a good fit between the model and the observed data (χ² = 0.046; P = 0.831; CFI = 1.000; TLI = 1.080; RMSEA < 0.001) and also adequate statistical power (0.927). Soil macronutrients had a significant direct positive effect on insect gall richness (Table 1), which was influenced by the positive effect of Ca and Mg on the insect gall richness (Appendix 3). On the other hand, both plant abundance and plant height had only indirect positive effects on insect gall richness via an increase in plant species richness (Figure 1; Table 1).
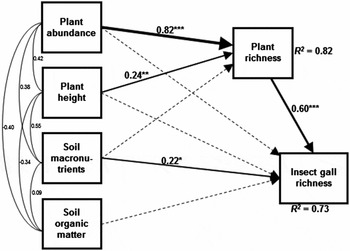
Figure 1. Path analysis for insect gall richness explained by plant abundance, plant height, soil macronutrients, soil organic matter and plant species richness of forty-nine 100-m2 savanna plots in Parque Nacional das Emas, Brazil. Numbers associated with paths between variables are path coefficients presented as standardized values (scaled by the standard deviations of the variables). Black arrows represent significant effects (*P < 0.05; **P < 0.01; ***P < 0.001) and dashed arrows represent non-significant effects (P > 0.05). The thickness of lines and arrows is proportional to effect size. Soil data obtained from Zava & Cianciaruso (Reference ZAVA and CIANCIARUSO2014).
Table 1. Explanatory model for insect gall richness (according to the path analysis in Figure 1) in forty-nine 100-m2 savanna plots in Parque Nacional das Emas, Brazil. Only variables with significant effects are presented (P < 0.05). Soil data obtained from Zava & Cianciaruso (Reference ZAVA and CIANCIARUSO2014).

DISCUSSION
The present study showed that both plant-related and environment-related factors are important to insect gall richness in a Neotropical savanna. The results indicate that the richness of galling insects is influenced by variables of vegetation structure, such as plant abundance and plant height, but only indirectly through the effect of these on plant species richness. The lack of any effect of vegetation structure on insect gall richness in addition to that mediated by plant species richness corroborates previous studies that suggested that the number of host plant species plays an important role in the maintenance of galling populations (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013, Cuevas-Reyes et al. Reference CUEVAS-REYES, QUESADA, HANSON, DIRZO and OYAMA2004).
The plant species richness hypothesis is one of the most tested and corroborated hypotheses for explaining the richness of phytophagous insects (review in Araújo Reference ARAÚJO2013). This hypothesis predicts a positive relationship between plant richness and galling insect richness, which can be explained by the fact that each plant species represents a potential niche for a galling species (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013). This positive correlation can be interpreted mathematically as meaning that more plant species are more likely to harbour more herbivores (Araújo Reference ARAÚJO2013). Host plant richness can influence both bottom-up processes acting on the galling insects, such as increasing the availability of a resource (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013), and top-down processes, such as decreasing parasitoid abundance and predation risk (Denno et al. Reference DENNO, GRATTON, DÖBEL and FINKE2003, Gámez-Virués et al. Reference GÁMEZ-VIRUÉS, GURR, RAMAN and NICOL2010).
There was no evidence that plots with greater plant abundance or larger plants hosted more insect galls. These results are contrary to the expectation of the resource concentration hypothesis (Root Reference ROOT1973), which predicts that insect gall richness would increase with increment in the plant abundance. Studies corroborating this hypothesis generally indicate that plant abundance dilutes predator pressure on aggregated plants (dilution effect) and acts as an indicator of abundance or nutritional quality of the host plant (Cuevas-Reyes et al. Reference CUEVAS-REYES, QUESADA, HANSON, DIRZO and OYAMA2004, Gonçalves-Alvim & Fernandes Reference GONÇALVES-ALVIM and FERNANDES2001). Nevertheless, no effect of plant abundance and height on insect gall richness was observed in studies where the effect of plant species richness was previously controlled (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013). In the present study only indirect effects of vegetation structure on insect gall richness were found, which are mediated by plant species richness. These findings suggest that previous studies that found positive effects of variables related to vegetation structure on richness of galling insects (Cuevas-Reyes et al. Reference CUEVAS-REYES, QUESADA, HANSON, DIRZO and OYAMA2004, Gonçalves-Alvim & Fernandes Reference GONÇALVES-ALVIM and FERNANDES2001) probably neglected the possibility that plant species richness can mediate these effects.
The path model showed that soil macronutrients have a direct positive effect on insect gall richness, after controlling for the possible indirect effects of soil characteristics via plant species richness. These results counter previous studies that corroborate the hypothesis that galling insect richness is negatively influenced by soil fertility in tropical environments (Fernandes & Price Reference FERNANDES and PRICE1988). Gonçalves-Alvim & Fernandes (Reference GONÇALVES-ALVIM and FERNANDES2001) investigated the effects of soil nutrients on galling richness in Brazilian savannas, and found high values of Mg (22.4 ± 25.0 cmol dm−3) and low levels of OM (0.25 ± 0.22 g kg−1) when compared with the PNE (Mg = 0.05 ± 0.01 cmol dm−3; OM = 42.4 ± 12.6 g kg−1). They found that these soil characteristics were negatively correlated with the insect gall richness. In another study, Cuevas-Reyes et al. (Reference CUEVAS-REYES, SIEBE, MARTÍNEZ-RAMOS and OYAMA2003) found lower insect gall occurrence in habitats with more fertile soils (in terms of amount of P) in Mexican tropical rain forests. Cuevas-Reyes et al. (Reference CUEVAS-REYES, QUESADA, HANSON, DIRZO and OYAMA2004) found that plants under poor soil conditions accumulate higher concentrations of secondary metabolites (e.g. phenols and terpenoids), which can be sequestered by galling insects during gall formation as a mechanism to protect them against natural enemies, resulting in a negative effect of soil fertility on insect gall distribution. Additionally, the inverse relationship between soil fertility and insect gall richness also may be due to the effect of soil fertility acting on plant species richness, as previously investigated by Blanche & Westoby (Reference BLANCHE and WESTOBY1995) in Australian forests. Since there was no effect of soil fertility on plant species richness in the present study, which may be explained by the fire regime in the PNE (Silva et al. Reference SILVA, BATALHA and CIANCIARUSO2013), it may be that unexplored factors that are not related to plant species richness may be acting.
I hypothesize that two plant factors, not related to plant species richness, can explain the positive direct effect of soil fertility on insect gall distribution: plant tissue quality and functional trait diversity. Soil characteristics may alter the nutritional quality of plant tissues (De Bruyn et al. Reference DE BRUYN, SCHEIRS and VERHAGEN2002), and there is much evidence that points to insect herbivores preferring plants with more nutritive tissues (review in Cornelissen et al. Reference CORNELISSEN, FERNANDES and VASCONCELLOS-NETO2008). Thus, high soil fertility possibly intensifies the occurrence of plants with more nutritive and vigorous modules (review in Altieri & Nicholls Reference ALTIERI and NICHOLLS2003), which favours the occurrence of a high density and richness of galling herbivores (Price Reference PRICE1991). Furthermore, soil fertility can also increase insect gall richness by influencing the functional traits of host plants. The soils of Brazilian savannas are characterized by high levels of acidity and low concentration of nutrients (Stark Reference STARK1970). These characteristics make it difficult for plants to recover nutrients lost to consumption by herbivores, and consequently select similar structural defensive traits of the plant species occurring in these habitats (Silva et al. Reference SILVA, BATALHA and CIANCIARUSO2013, Zava & Cianciaruso Reference ZAVA and CIANCIARUSO2014). Thus, on sites with higher soil fertility, these pressures for homogenization of plant traits are reduced, which can lead to greater diversity of traits to be exploited by insect galling. Further investigations are needed to test if soil fertility influences resource quality and functional trait diversity of host plants, and how these affect the richness of insect galls.
This study showed that plant species richness has an important influence on insect gall richness in Neotropical savannas at the local level, and complements studies that have shown this pattern at the regional level (Araújo et al. Reference ARAÚJO, SCARELI-SANTOS, GUILHERME and CUEVAS-REYES2013, Reference ARAÚJO, CUEVAS-REYES and GUILHERME2014). These findings represent a major advance from previous studies because it is the first study to identify the effects of vegetation structure and soil characteristics on insect gall richness in Neotropical savannas, while controlling for their indirect effects via plant species richness. The descriptors of vegetation structure (i.e. plant abundance and plant height), were found to influence insect gall richness, but only indirectly via increasing plant species richness. Thus, future studies that investigate the influence of vegetation-related factors on the diversity of gall-inducing insects should take into account the importance of plant species richness.
ACKNOWLEDGEMENTS
I am very grateful to Fernando Sobral, Leonardo Bergamini and Leandro Maracahipes for help in field surveys; to Paulo Zava and Marcus Cianciaruso for providing soil data; and to Paulo Corgosinho and Erik Wild for suggestions and English revision of the manuscript.
Appendix 1. Data summary for plant characteristics (richness, abundance and height), soil characteristics (Ca, Mg, P, N and OM) and insect gall richness of forty-nine 100-m2 savanna plots in Parque Nacional das Emas, Brazil. Soil data obtained from Zava & Cianciaruso (Reference ZAVA and CIANCIARUSO2014). Values of plant height and soil characteristics represent the mean values for each plot.

Appendix 2. Pearson's correlation table for plant height, plant abundance, plant richness and soil characteristics (Ca, Mg, P, OM and N) of forty-nine 100-m2 savanna plots in Parque Nacional das Emas, Brazil. Values marked with asterisks are statistically significant (*P < 0.05; **P < 0.01; ***P < 0.001). Soil data obtained from Zava & Cianciaruso (Reference ZAVA and CIANCIARUSO2014).

Appendix 3. Pearson's correlation table of soil characteristics with insect gall richness of forty-nine 100-m2 savanna plots in Parque Nacional das Emas, Brazil. Values marked with asterisks are statistically significant (*P < 0.05; **P < 0.01; ***P < 0.001). Soil data obtained from Zava & Cianciaruso (Reference ZAVA and CIANCIARUSO2014).






