


Introduction
The processes and events associated with the cessation of the building and use of the megalithic tombs at Newgrange, one of Europe’s most renowned archaeological sites, remain imperfectly understood. The largest of the Neolithic tombs at Newgrange was constructed c. 3300–3000 cal BC (O’Kelly Reference O’Kelly1982; Lynch et al. Reference Lynch, McCormick, Shee Twohig, McClatchie, Molloy, Schulting, OCarroll and Sternke2014) and is widely regarded as a key focal point in the archaeologically rich Brú na Bóinne UNESCO World Heritage complex, Co. Meath, Ireland (eg Stout Reference Stout2002; Smyth et al. Reference Smyth, Brady, Chadwick, Condit, Cooney, Doyle, Guinan, Potterton, Stout and Tuffy2009; Hensey Reference Hensey2015) (Figure 1). The tomb’s entrance is oriented such that at sunrise, for a small number of days around the winter solstice, a shaft of light enters through a portal above the long entrance passage, illuminating the interior of the tomb (Patrick Reference Patrick1974; O’Kelly Reference O’Kelly1982; Prendergast Reference Prendergast1991; Hensey Reference Hensey, Papadopoulos and Moyes2017; Williams Reference Williams2019). This annual event was a key concern of those who constructed the monument, as evidenced by an evolving sequence of rebuilding (c. 3305–3020 cal BC; O’Kelly Reference O’Kelly1982; Eriksen Reference Eriksen2008; Lynch et al. Reference Lynch, McCormick, Shee Twohig, McClatchie, Molloy, Schulting, OCarroll and Sternke2014; Hensey Reference Hensey2015) that sought to emphasise and draw meaning from specific elements of the site’s interior and exterior features (eg centring on themes of lightness and darkness; for review, see Hensey Reference Hensey2015).
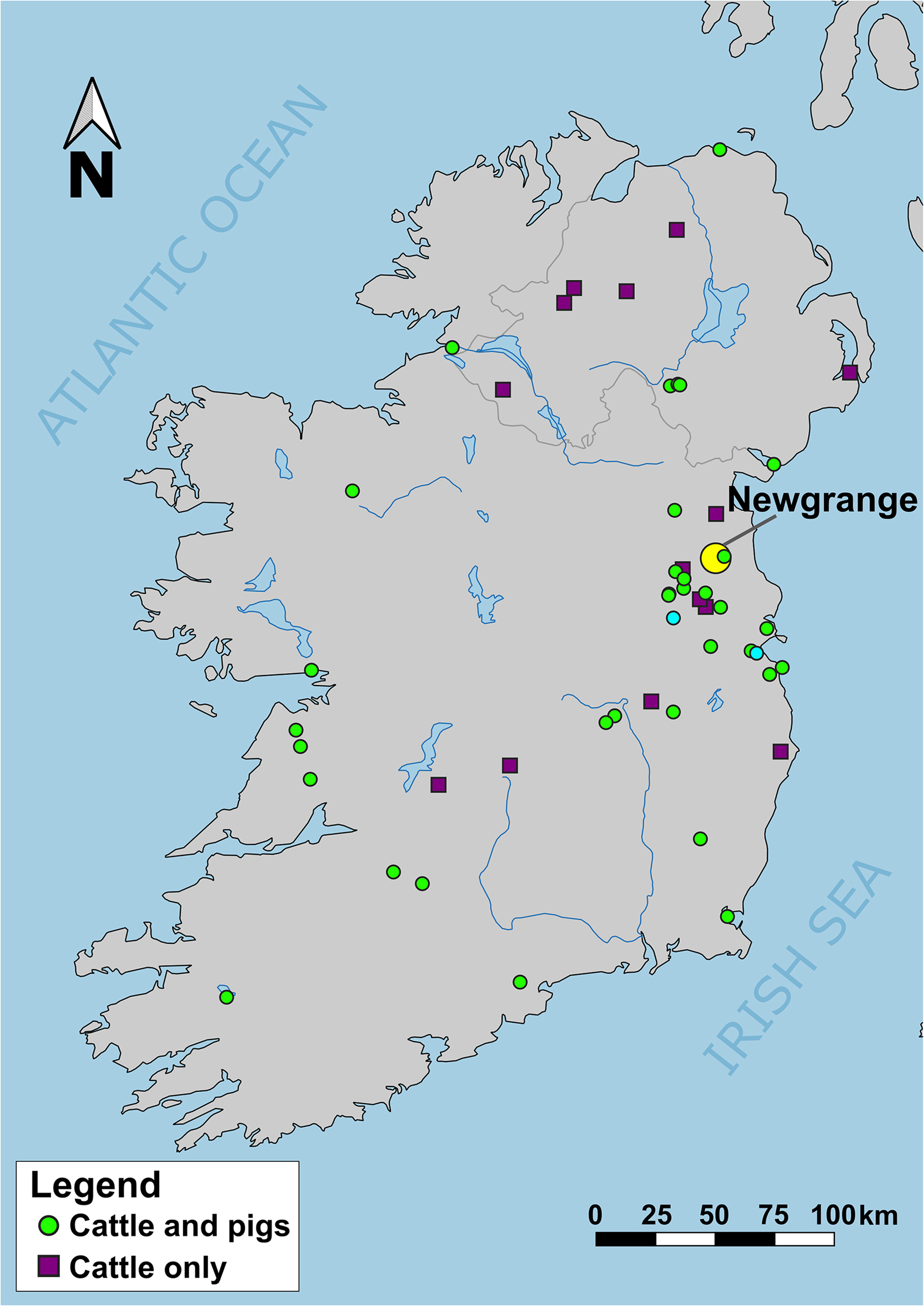
Figure 1. Map showing location of Newgrange (large yellow circle) relative to sites with cattle and pig baseline data, spanning from the Neolithic to post-medieval periods. See Figure S1 for version showing site names.
Despite these spectacular archaeological patterns, the circumstances surrounding the closure of the tomb c. 2900–2700 BC remain uncertain (for review, see Hensey Reference Hensey, Papadopoulos and Moyes2017), especially its relationship with the activity outside it. We know, for instance, that people continued utilising the space around the tomb during the 3rd millennium BC and the 1st millennium AD. The faunal assemblage excavated from this area is remarkable, representing a rare occurrence of a large body of evidence for human–animal relationships in Ireland, but the dating and character of these deposits are poorly understood (O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983; Sweetman et al. Reference Sweetman, O’Sullivan, Gunn, Monk, Donnabh and Scannell1985; Reference Sweetman, McCormick and Mitchell1987; Carlin Reference Carlin2017; Reference Carlin2018). For instance, it is not known to what extent the central themes that guided the tomb’s earlier construction and use persisted or influenced the activities that generated these later deposits. Did those who inhabited the Brú na Bóinne landscape continue to organise some of their activities around the winter solstice? And if so, what might this say about continuity of these cultural concepts through time? How can the large faunal assemblage inform us about this?
We analysed age-at-slaughter, isotope compositions and radiocarbon dates for pig remains excavated around the periphery of the monument to assess the nature and seasonality of activity in the centuries that followed the construction, use, and sealing of the large tomb at Newgrange, and the implications these have for the wider Brú na Bóinne landscape. We find that the large volume of animal bone distributed across the site, which is thought to reflect feasting activities, likely dates to c. 2600–2450 cal BC, generally regarded as the end of the Neolithic, just prior to the Chalcolithic. Furthermore, the unique diets of pigs, and the ages at which they were typically slaughtered, suggest they were processed and consumed around the time of the winter solstice. These data suggest that Newgrange remained a focus of attention during the winter solstice, even after the interior of the tomb was sealed off (sometime after c. 2900–2700 BC) and we explore the potential implications of this.
Context
A brief outline of Newgrange in the deep past
O’Kelly’s 1962–1975 excavations at Newgrange revealed the large mound to be a Neolithic developed passage tomb with a cruciform arrangement of chambers and elongated entrance passage (O’Kelly Reference O’Kelly, Daniel and Kjaerum1973; Reference O’Kelly1982; O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983; Figure 2). Faunal and human remains were found in the chambers, along with objects typical of Irish passage tombs, however these had been highly disturbed in recent centuries (O’Kelly Reference O’Kelly1982; Smyth et al. Reference Smyth, Geber, Carlin, O’Sullivan, Griffiths, Hofmann, Cummings, Bjørnevad-Ahlqvist and Iversen2025).
The chronology of the tomb and activity within the broader Newgrange complex have been poorly understood due to a limited number of radiocarbon dates and significant misunderstandings from earlier interpretations. However, a clearer picture of the multi-period nature of the site is beginning to emerge. We provide a brief outline of this as context for our results.
Fitting firmly within the by now well-established chronological horizon of developed passage tombs (eg Schulting et al. Reference Schulting, Bronk Ramsey, Reimer, Eogan, Cleary, Cooney and Sheridan2017a; Reference Schulting, McClatchie, Sheridan, McLaughlin, Barratt and Whitehouse2017b) the large Newgrange tomb was built episodically, and initially used between c. 3300–3000 cal BC, but deposition within it continued until c. 2700 cal BC, when its interior seems to have been sealed off (O’Kelly Reference O’Kelly1982; Lynch et al. Reference Lynch, McCormick, Shee Twohig, McClatchie, Molloy, Schulting, OCarroll and Sternke2014; Smyth et al. Reference Smyth, Geber, Carlin, O’Sullivan, Griffiths, Hofmann, Cummings, Bjørnevad-Ahlqvist and Iversen2025; Carlin et al. Reference Carlin, Griffiths, Kanne and Smythforthcoming).
Various additions continued to be made to the monument (Hensey Reference Hensey2015) during its initial use including the encircling of the large tomb’s kerbed perimeter with the ‘great stone circle’, a ring of standing stones — some of which were decorated with megalithic art (Williams Reference Williams2023). Encircling the front of the tomb was an extensive wedge-shaped layer of quartz and granite stones. Subsequently, at the south-eastern limit of this layer, a large oval enclosure of post-pits was created c. 2700–2400 BC which encircled the Site Z passage tomb. Excavations revealed that these pits contained animal bones (mainly cattle and pig) and Grooved Ware pottery. Later Grooved Ware-associated activity was conducted around the frontal perimeter of the large passage tomb between 2700–2450 cal BC, including the creation of several stone-lined hearths and numerous pits and postholes (for overview, see Carlin Reference Carlin2017).
Subsequently, an earth and stone layer (labelled the ‘Beaker layers’; O’Kelly Reference O’Kelly1982; O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983) formed on top of the exterior features. This comprised chronologically mixed deposits containing large quantities of lithics, animal bone, and ceramics including Grooved Ware and Beaker pottery, and other materials dating between the 4th millennium BC and the 4th century AD. The chronological insecurity of these ‘Beaker layers’ is exemplified by their apparent occurrence both under and over a bank of stoneless yellow boulder clay that flanked the tomb’s outer perimeter from its entrance to beyond the western limit of excavation, and which sealed some of the stone hearths. Known as the Yellow Clay Bank (hereafter YCB; Figure 2), the stratigraphy and temporal attribution of this feature has sparked considerable debate (O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983; Mount Reference Mount1994; Cooney Reference Cooney2006; Carlin Reference Carlin2017).

Figure 2. Newgrange site plan (after Lynch et al. Reference Lynch, McCormick, Shee Twohig, McClatchie, Molloy, Schulting, OCarroll and Sternke2014, fig. 2) showing location of eastern (red) and western (green) excavation units (approximate, after O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983, fig. 2).
O’Kelly (Reference O’Kelly1982; O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983) considered the ‘Beaker layers’ to represent about 500 years of Beaker-period occupation which began c. 2500 BC at the start of an economic collapse that immediately followed the peak of the passage tomb related activity. He used the term ‘Late Neolithic/Beaker period’ to characterise what he regarded as evidence for both continuity and change between the passage tomb and Beaker-associated activities at the site. More recently, there have been substantial critiques of this, especially because activity associated with Grooved Ware during the earlier 3rd millennium BC was not fully recognised by O’Kelly (Carlin Reference Carlin2017). It is now understood that the ‘Beaker layers’ represent a long sequence of depositional activity from 3300–2000 BC and again from the 1st to 4th centuries AD, strongly focused on the area immediately outside the tomb entrance (for overview, see Carlin Reference Carlin2017; Reference Carlin2018). Notably, 151 horse bones and teeth occurred within the ‘Beaker layers’ but two of these teeth have been radiocarbon dated to the start of the 1st millennium AD (Bendrey et al. Reference Bendrey, Thorpe, Outram and Van Wijngaarden-Bakker2013). These dates support wider concerns about the potential for admixture of artefacts from the later half of the 3rd millennium BC with earlier and later materials (Carlin Reference Carlin2018). Significantly, no horse remains occurred under the YCB, suggesting that the deposits beneath it are predominantly from the 3rd millennium BC, while at least some of the deposits above it were still being formed around AD 100. This deposition ceased in the 4th–5th century AD, and evidence for further activity is lacking until the tomb was re-opened in AD 1699.
An enigmatic faunal assemblage
The horse bones formed part of a much larger faunal assemblage that was recovered from the chronologically mixed ‘Beaker layers’ (O’Kelly Reference O’Kelly, Daniel and Kjaerum1973; Reference O’Kelly1982; O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983). Zooarchaeological analyses by Van Wijngaarden-Bakker (Reference Van Wijngaarden-Bakker1974; Reference Van Wijngaarden-Bakker1986) identified more than 12,000 elements and her interpretations adhered to the excavator’s view (O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983) that these belonged to the same chronological horizon and represented food remains from ‘domestic’ activities at a Beaker settlement. However, aspects of these interpretations have been questioned as information has emerged about the multi-period nature of the deposits (Carlin Reference Carlin2017; Reference Carlin2018). The anachronistic radiocarbon dates from horses (Bendrey et al. Reference Bendrey, Thorpe, Outram and Van Wijngaarden-Bakker2013) have led to the recognition that the contemporaneity of the faunal assemblage with Beaker-associated activities is less certain than originally thought.
Other excavations outside the rear and side of the larger passage tomb produced very few animal bones (Ó Ríordáin & Ó hEochaidhe Reference Ó Ríordáin and Ó hEochaidhe1956; O’Kelly Reference O’Kelly1982; Lynch et al. Reference Lynch, McCormick, Shee Twohig, McClatchie, Molloy, Schulting, OCarroll and Sternke2014) thereby indicating that faunal remains at Newgrange were predominantly deposited at the front of the monument. Indeed, the greatest frequency of bones and individual animals were identified from the deposits nearest the entrance. Unsurprisingly, there has been significant debate regarding whether these faunal remains reflect ceremonial or settlement activities, or some combination of the two (eg Mount Reference Mount1994; Cooney Reference Cooney2006; Lynch et al. Reference Lynch, McCormick, Shee Twohig, McClatchie, Molloy, Schulting, OCarroll and Sternke2014; Carlin Reference Carlin2018). While Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1986) interpretations were hindered by taphonomic factors, including extensive erosion of the bone surfaces, she generated valuable insights into how people interacted with these animals, particularly pigs. The assemblage was dominated by pig and cattle remains, both composed of particularly large individuals. Bones found in association suggested that specific portions of pig carcasses (notable body portion units included the thoracic cage as well as individual limbs) were systematically disposed of while still articulated, often in concentrations and suggesting specialised activity areas (Appendix S1). Furthermore, the recovery of intact bones showed that marrow extraction had not been systematically performed. Together these patterns were taken as evidence for on-site slaughter, butchery, and preservation by smoking to produce stable pork products.
Van Wijngaarden-Bakker also examined seasonality of pig slaughter based on wear stages of third molars. Unfortunately, her report did not include all toothwear data and as such, it is unclear if the age estimates accurately reflect the population demographics or if, as is implied in the text, they reflect a subset that was selected to examine seasonality. Despite these ambiguities, the seasonality data are of significant value. Assuming prehistoric pig oestrous cycles were similar to wild pigs today, Van Wijngaarden-Bakker (Reference Van Wijngaarden-Bakker1986) supposed birth would be largely constrained to May. She found that 68 % of pigs were killed between October and April of their second (33 %) or third (35 %) years of life.
Based on patterns in epiphyseal fusion, Van Wijngaarden-Bakker found that 87 % of pigs were older than two years at death and a majority (57 %) were killed between the ages of 2 and 2.5 years, which she suggested showed a peak of slaughter between May and November of their third year. Although the fusion and tooth data indicate only a small overlap in October and November, she argued that these data corroborated one another, and that most pig slaughter was confined to the autumn–winter (November to April) season. In fact, her interpretation of the fusion data indicated predominantly summer slaughter while her tooth data indicated predominantly winter slaughter.
Despite the interpretive challenges, a cohesive narrative emerged from Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1986) analyses: large pigs were slaughtered in the vicinity of the monument entrance in the autumn–winter months and their bodies were divided into portions that were used at different locations across the site (Appendix S1). These activities, she supposed, were dedicated to producing fresh and preserved pork products for victualing the local settlement and exchange with neighbours for other resources. Cooney (1987; Reference Cooney1991) and Mount (Reference Mount1994) noted that these findings could be interpreted differently if the original settlement interpretation of the site is reconsidered. Specifically, they question Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1986) attribution of the faunal assemblage to ‘domestic’ activities and instead suggest these taxonomic, age, and spatial patterns are more parsimoniously interpreted as reflecting ceremonial activities (Appendix S1).
McCormick (Reference McCormick, Murphy and Whitehouse2007) added, based on the widely accepted association between pig keeping and pannage-focused husbandry in woodlands (ie use of acorns and other mast to feed pigs; see below) that the high proportion of pigs may reflect increasing oak and other woodland cover during the 3rd millennium BC. While reviews of the Irish palaeoecological evidence reveal broader-scale trends for the ebb and flow of woodland cover in the prehistoric period (Plunkett Reference Plunkett1999; O’Connell & Molloy Reference O’Connell and Molloy2001; McClatchie et al. Reference McClatchie, Bogaard, Colledge, Whitehouse, Schulting, Barratt and McLaughlin2014; Whitehouse et al. Reference Whitehouse, Schulting, McClatchie, Barratt, McLaughlin, Bogaard, Colledge, Marchant, Gaffrey and Bunting2014), specific evidence for the 3rd millennium BC remains patchy, making this difficult to evaluate (O’Connell & Molloy Reference O’Connell and Molloy2001; McClatchie & Potito Reference McClatchie and Potito2020; Appendix S2).
These ongoing debates notwithstanding, prehistoric faunal assemblages of the size excavated at Newgrange remain extremely rare across Ireland (McCormick Reference McCormick, Murphy and Whitehouse2007; McClatchie et al. Reference McClatchie, Schulting, McLaughlin, Colledge, Bogaard, Barratt and Whitehouse2022; Stout et al. Reference Stout, Stout, Beglane, Brady, Davis and Leigh2023). Establishing the timing and nature of human–animal relationships at this site is therefore of significance beyond Newgrange.
The isotopic ecology of Irish pig husbandry
Stable carbon (δ 13C) and nitrogen (δ 15N) isotope analysis of bone collagen offers a well-established method for reconstructing both human and animal diets in the past (Ambrose Reference Ambrose1990). At a molecular level, materials from foods eaten are directly incorporated into our tissues and consumption of isotopically distinctive foods can be detected in archaeological bones and teeth (Appendix S3).
Holocene-scale surveys of Irish livestock, coupled with knowledge of global axes of variation in terrestrial isotope ecology (Guiry et al. Reference Guiry, Beglane, Szpak, Schulting, McCormick and Richards2018; Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a), provide a robust interpretive framework for exploring human–pig–environment relationships at Newgrange. As Ireland has a C3-dominated environment, there are two main factors at play when interpreting livestock δ 13C. First, feeding in open (eg meadows or cleared fields) versus closed (eg dense woodland) environments will result in animals with higher and lower δ 13C values, respectively (Van der Merwe & Medina Reference Van der Merwe and Medina1991). This relationship is widely recognised in archaeological research on European fauna (eg Noe-Nygaard et al. Reference Noe-Nygaard, Price and Hede2005; Drucker et al. Reference Drucker, Bridault, Hobson, Szuma and Bocherens2008; Lynch et al. Reference Lynch, Hamilton and Hedges2008) and is the basis of a recent exploration of human–cattle relationships, which included samples from Newgrange, showing that cattle from the site were typically maintained in denser woodland environments compared to cattle husbanded in other time periods (Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a).
Second, animal diets targeting photosynthetic (ie leaves and foliage) and non-photosynthetic (eg mast, including acorns and other nuts) portions of plants will have lower and higher δ 13C values, respectively (Craig Reference Craig1953). While this relationship has not been utilised for interpretations of archaeological pig husbandry in Europe, it has been demonstrated by ecological research on plant tissues (Cernusak et al. Reference Cernusak, Tcherkez, Keitel, Cornwell, Santiago, Knohl, Barbour, Williams, Reich and Ellsworth2009), as well as hair and muscle samples from woodland animals in other regions (eg Sarà & Sarà Reference Sarà and Sarà2007; Selva et al. Reference Selva, Hobson, Cortés-Avizanda, Zalewski and Donázar2012; Pauli et al. Reference Pauli and Steffan2019; Karmanova et al. Reference Karmanova, Feoktistova and Tiunov2023). Archaeological isotopic research surveying ecologically diverse woodland fauna has confirmed that mast feeding specialists have higher δ 13C values relative to their non-mast specialist counterparts (Guiry et al. Reference Guiry, Orchard, Royle, Cheung and Yang2020; Reference Guiry, Orchard, Needs-Howarth and Szpak2021). In the context of livestock at Newgrange, and indeed all European archaeological sites where cattle and pigs occur together, there is an important distinction to be made. While oak mast (acorns), a primary component of mast in many European woodlands, is poisonous in large quantities for cattle, it is sought out by pigs (this is pannage feeding: Szabó Reference Szabó and Rotherham2013; Wealleans Reference Wealleans2013). With respect to δ 13C, therefore, strongly woodland-oriented husbandry will push cattle and pig δ 13C signals in opposite directions. For cattle, foraging in a closed canopy woodland will result in lower δ 13C, and for pigs, consuming large quantities of tree mast, even in those same closed environments, will result in higher δ 13C.
With respect to δ 15N, which undergoes a stepwise increase between trophic levels, there are three primary axes of variation for Irish pigs. First, as omnivores, they can consume animal products, meaning that pig δ 15N will vary along a herbivory-to-carnivory spectrum, something which can inform on a variety of husbandry practices (eg use of food scraps to raise pigs: DeNiro & Epstein Reference DeNiro and Epstein1981; Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). Sources of animal protein can include meat, dairy byproducts, and faeces. Second, milk-feeding, which effectively sets the feeder a trophic level above the mother, means that infant and recently weaned animals will also show higher δ 15N (Fogel et al. Reference Fogel, Tuross and Owsley1989). Third, research exploring wider trends in Irish livestock δ 15N (Guiry et al. Reference Guiry, Beglane, Szpak, Schulting, McCormick and Richards2018) has highlighted the importance of isotopic baseline shifts driven by human land management. While these shifts, involving increases both in average baseline δ 15N and overall herbivore δ 15N variation, primarily begin in the Middle to Late Bronze Age, and are therefore not relevant to our analysis of human–pig–environment relationships at Newgrange, they are relevant when we consider Newgrange pig diets relative to those in later time periods.
In addition, Newgrange is situated on the River Boyne, providing access to freshwater and diadromous (marine) fish. This means that pigs may have had opportunities to consume foods linked to a wider range of carbon and nitrogen sources than would be expected for local terrestrial foods (Guiry Reference Guiry2019). Further, as aquatic ecosystems have more trophic levels than terrestrial ecosystems (Minagawa & Wada Reference Minagawa and Wada1984), pigs at Newgrange may also have accessed foods with higher δ 15N values than expected for the local terrestrial food web (Guiry et al. Reference Guiry, Noël, Tourigny and Grimes2012; 2023b).
The isotopic compositions of bone collagen reflect a complex mixture of biogeochemical processes. For this reason, best practice is to contextualise interpretations within a framework of data from other fauna from the same site and time period, and from the wider region (Katzenberg Reference Katzenberg1989). In this study we have robust baselines composed of herbivorous cattle from the same site and timeframe (Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a) and of pigs spanning much of the Holocene (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). We acknowledge that cattle and pigs have differing digestive physiologies, but do not consider this to be an interpretive issue for comparing isotopic compositions in this study (see Appendix S3).
Research questions and hypotheses
We conducted these analyses as part of a larger project aimed at uncovering human–animal–environmental relationships throughout the Holocene (Guiry et al. Reference Guiry, Beglane, Szpak, Schulting, McCormick and Richards2018; Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a; 2025). Within the context of that work, we recognised that isotopic compositions and associated radiocarbon dates from pigs at Newgrange represented a special case meriting closer consideration. We elected to interpret these data separately from the main body of Irish pigs (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025) and bring them to bear on questions generated by the debate that has stemmed from Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1974; Reference Van Wijngaarden-Bakker1986) work.
These research questions (RQs) fall under two themes.
Theme 1, Chronology. We use radiocarbon dating to ask:
-
1. Are these pigs contemporaneous with Beaker pottery in Ireland (c. 2450–2050 BC; Carlin Reference Carlin2018)?
-
2. If not, what temporal scale do we see represented?
-
3. Does the way humans and pigs interacted change over time?
Theme 2, Husbandry. We use isotope analyses and re-examination of pig ageing data to ask:
-
4. Were the pigs raised in a woodland-oriented (ie pannage-focused) husbandry regime?
-
5. If so, what are the implications for our understanding of seasonality of the assemblage at Newgrange?
-
6. More broadly, how do husbandry practices compare with other places and times?
Materials and methods
Samples came from the western and eastern units (Figure 2). Samples from the western unit can be further divided as coming from above and below the YCB, a stratigraphic feature which hemmed the kerbstones extending out from the monument a distance of 5–7 m (Figure 2; see Appendix S4).
We performed δ 13C and δ 15N analyses on bone collagen extracted from 69 pigs (a fraction of the 4208 pig bones recovered; Van Wijngaarden-Bakker Reference Van Wijngaarden-Bakker1986) including 61 from the western unit (25 above and 36 below the YCB, respectively) and eight from the eastern unit. Collagen extraction and purification, as well as isotope ratio mass spectrometry analyses and calibration followed well-established protocols (Appendix S5, Tables S3–6). For δ 13C and δ 15N, standard uncertainties were ±0.14 ‰ and ±0.19 ‰ (Szpak et al. Reference Szpak, Metcalfe and Macdonald2017). Quality control used carbon (>13.8 %) and nitrogen (>4.0 %) concentrations (Ambrose Reference Ambrose1990) and conservative C:N criteria, tolerating δ 13C shifts no greater than –0.5 ‰ (Guiry & Szpak Reference Guiry and Szpak2021). Statistical comparisons were performed using PAST version 4.13 (Hammer et al. Reference Hammer, Harper and Ryan2001; Appendix S5).
We reviewed Van Wijngaarden-Bakker’s interpretation of pig ageing and seasonality using now standard age categories and methodologies (Appendix S6, Tables S7-9).
We radiocarbon dated four pig bones from the western unit, two each from above and below the YCB, and three from the eastern unit. To this we can add three published dates on cattle calcanei, one from the eastern unit, and one each from the western unit above and below the YCB (Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a; Table S2). All 10 samples represent discrete individuals. Dates were calibrated using IntCal20 (Reimer et al. Reference Reimer and Talamo2020) and modelled using OxCal v.4.4.4 (Bronk Ramsey Reference Bronk Ramsey2021) in four ways.
Firstly, the dated remains were included in two simple models to estimate the duration of depositional activity represented by the bones:
-
1) Most conservatively, all 10 animal bone dates were included in a single bounded phase and OxCal was used to estimate start date, end date, and duration (‘Model 1’).
-
2) In response to observable clustering of pig dates, and to explore human–pig interactions at Newgrange specifically, this model was repeated for these seven samples alone to assess the possibility that the abundance of lightly fragmented, anatomically grouped pig remains might represent a more constrained episode of activity within a wider phase of site use (‘Model 2’).
Exploring Model 2 further, we conducted simulations using the ‘uncalibrate’ function in rcarbon v.1.5.1 (Crema & Bevan Reference Crema and Bevan2021) in R v.4.3.2 (R Core Team 2023) to assess the feasibility of the observed pig results under the extreme hypothesis of a single-year deposition event. To assess species-specific difference in temporality while removing the effects of differing sample sizes, we also compared (a) all three cattle dates modelled as a single phase, with (b) all possible combinations of three pig dates, each modelled in the same way.
Additional models explored the relationship between the pig bone deposits and the YCB under differing assumptions regarding the integrity of the overlying deposits.
-
3) ‘Model 3’, a tentative stratigraphic model, is based on the assumption (following O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983) that the bank was part of the same overall phase of activity as the bone deposits and that the overlying pig bones were not redeposited, allowing a tight estimate to be made for the date of construction of the YCB. The eastern and western units were treated as separate, parallel entities within an overall bounded phase, and the western unit modelled as a sequence with samples from below and above the YCB separately. The cattle dates were treated as termini post quos using the ‘After’ function in OxCal to reflect the possibility of reworking.
-
4) In contrast, ‘Model 4’ allowed for the possibility that the pig samples from above the YCB represent materials in secondary context, as highlighted in recent reconsideration of the YCB stratigraphy (see below). All pig dates were included in a single bounded phase (as in Model 2) on the assumption that their initial deposition represented a single phase of activity, even if some may have been subsequently redeposited. The ‘Last’ function in OxCal was then used to estimate the probability distribution of the latest out of (a) the two modelled pig dates from under the YCB (UOC-16271, UOC-16272), and (b) the single cattle date from under the YCB (UOC-16270, itself not stratigraphically modelled). This estimated date distribution is then taken as the terminus post quem for the construction of the YCB.
Details, as well as OxCal code for all models and R code for simulations, are provided in Appendix S15.
Results
Isotope analysis
Most pigs (n = 67, 97 %) produced elemental concentrations and C:N ratios meeting quality control criteria (Figure 3; Table S1). Pigs from the western (n = 59) and eastern (n = 8) units produced mean δ 13C values of –21.5±0.3 ‰ and –21.6±0.3 ‰, respectively, and δ 15N values of +5.6±0.3 ‰ and +5.9±0.5 ‰, respectively (note all isotope compositions are quoted at one standard deviation). Within the western unit, pigs from above (n = 25) and below (n = 34) the YCB produced mean δ 13C values of –21.6±0.3 ‰ and –21.5±0.3 ‰, respectively, and δ 15N values of +5.7±0.6 ‰ and +5.5±0.6 ‰, respectively. Statistical comparisons showed no significant differences for δ 13C and δ 15N values compared between eastern and western units and subsamples from the western unit (ie above and below the YCB versus one another and each of these western unit subsamples versus the eastern unit sample; Appendix S7).

Figure 3. Isotopic compositions of Newgrange pigs (coloured symbols) in context of all published archaeological pig data from Ireland (hollow grey circles; Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). Newgrange samples are colour and shape coded by context: red square = eastern unit; yellow diamond = western unit, above the YCB; blue circle = western unit, below the YCB.
Compared with cattle (excluding bones that came from calves; n = 34) from Newgrange (Figure 4;), pigs (n = 67) have significantly higher δ 13C and lower δ 15N values. Compared with cattle from the early medieval period in Ireland (the period with the most 13C-enriched cattle, see below; n = 71) Newgrange pigs have significantly higher δ 13C values (Appendix S7).

Figure 4. Isotopic compositions of Newgrange pigs in the context of archaeological cattle from Ireland. A) Kernel density histograms for Newgrange pig δ 13C relative to key cattle comparison groups: contemporaneous cattle from Newgrange (NG) (orange), early medieval (EM) cattle (yellow; ie cattle with highest δ 13C for all time periods), and all cattle (light grey; n = 382). B) Newgrange pig δ 13C and δ 15N (dark grey) in context of all cattle (hollow light grey), with Newgrange cattle (orange), other Neolithic Early Bronze Age cattle (blue), and early medieval cattle (yellow). C) Panel B reproduced as standard ellipse areas (1σ) and convex hulls. D) Violin and box plots showing kernel density for δ 15N of both pigs (dark grey) and cattle (orange) from Newgrange, with cattle groups for Neolithic–Early Bronze Age (blue) and all cattle (light grey) shown for reference.
We also compared Newgrange pigs to pigs from other time periods across the Irish Holocene (Figure 5; see Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). Only samples that passed conservative quality control criteria, designed to ensure that potential humic acid contaminants do not shift δ13C values by more than –0.5 ‰, were included (Guiry & Szpak Reference Guiry and Szpak2021). Comparison groups (Figure 1; see Appendix S7 for full details) are the Neolithic and Early Bronze Age (c. 4000 to 1500 BC; n = 16), the Middle–Late Bronze Age and Iron Age (c. 1500 BC to AD 400; n = 119), and the medieval and post-medieval periods (c. AD 400 to 1900; n = 9). Compared with pigs from other Neolithic and Early Bronze Age sites, pigs from Newgrange have higher δ 13C (though not significantly) and higher δ 15N (significantly) values (Appendix S7). Relative to pigs from the rest of the Bronze Age and the Iron Age, Newgrange pigs have significantly higher δ 13C (by 0.7 ‰) and lower δ 15N (by 0.8 ‰). Similarly, compared with pigs from the medieval and post-medieval periods, pigs from Newgrange have significantly higher δ 13C (by 0.3 ‰) and lower δ 15N (by 2.7 ‰).

Figure 5. Isotopic compositions of Newgrange pigs in the context of all published archaeological pig data from Ireland. A) Kernel density histograms for Newgrange pig δ 13C (dark grey) relative to other periods: Neolithic–Early Bronze Age (blue, n = 16), Middle–Late Bronze Age and Iron Age (orange, n = 119), and the medieval and post-medieval periods (yellow, n = 103). B) Newgrange pig δ 13C and δ 15N in context of all pigs (same colour coding as Panel A). C) Panel B reproduced as standard ellipse areas (1σ) and convex hulls. For Panels A, B, and C, pig δ 13C for early medieval, later medieval, and post-medieval has been adjusted for trophic enrichment factor (by –0.20 ‰, –0.18 ‰, and –0.26 ‰, respectively, per Guiry et al. (Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). D) Violin and box plots showing kernel density for Newgrange pigs and baseline-corrected δ 15N from other time periods. Baselines for all later periods (ie M-LBA to PM) have been corrected to the earliest (Neo/EBA) period based on the difference between herbivore δ 15N in respective time periods (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025).
Given the known, spatially broad δ 15N baseline shifts occurring across Ireland in later prehistory, comparison of Newgrange pig δ 15N with results from other time periods requires small adjustments to correct mean pig isotopic compositions from later time periods for increasing baseline δ 15N (as outlined in Figure 5d; Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). Having already noted that the difference between cattle and pig means at Newgrange, though significant, is very small, the presence of significantly higher δ 15N values from Newgrange pigs relative to pigs from other Neolithic–Early Bronze Age sites should be considered carefully. We note that cattle at Newgrange also have higher δ 15N values (by c. 1.2 ‰) than cattle at other prehistoric sites (ie Neolithic and Early Bronze Age; n = 36 from 12 sites, mean = +5.1±0.7 ‰ (Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a)) suggesting local δ 15N baselines around Newgrange were higher than in other areas.
Radiocarbon dates
Simple OxCal modelling of all 10 dates together (seven new dates on pigs plus three on cattle from Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a; Figure 6: Model 1; Table S2) produced an estimated start date for this phase of activity of 2760–2480 cal BC (95 % probability; as are all date estimates quoted here) and an estimated end date of 2480–2300 cal BC, with a median estimated duration of 210 years (50–421 years; Figure 7: Model 1).

Figure 6. Probability distributions of dates on fauna from Newgrange. Each distribution represents the relative probability that an event occurs at a particular time. For each of the dates two distributions have been plotted: one in outline, which is the result of simple radiocarbon calibration, and a solid one, based on the chronological model used. Model 1 includes all dated pig (grey) and cattle (red) specimens as a single OxCal ‘phase’ with estimated start and end boundaries (blue). Model 2 repeats this with the cattle excluded.

Figure 7. Modelled durations of depositional activity for eastern and western units at Newgrange. Model 1: single phase with all 10 dates; Model 2: single phase with seven pig dates only.
Modelling only the seven pig samples as a single bounded phase (Figure 6: Model 2) gives a much shorter median estimated duration of 25 years (0–132 years; highest probability density at <10 years; Figure 7: Model 2). Start and end boundaries are very similar to each other, at 2600–2470 cal BC and 2570–2440 cal BC, respectively, although each has a double peak due to the shape of the calibration curve. Taken alone, the pig dates suggest a very brief period of depositional activity in either the mid–late 26th century or first half of the 25th century cal BC. The results of six of seven pig dates cluster more tightly than might be expected even in replicates on the same individual animal, given the error ranges inherent in dating. Simulation shows that this remarkable clustering is not implausible, however (Appendix S14): c. 15 % of sets of seven uncalibrated dates simulated from any given single calendar year within the scope of the study had a standard deviation below that observed here, and very similar results were obtained for hypothetical 10- or 20-year episodes of activity.
Contrasting with the tightly clustered pig dates, modeling the three cattle dates as a single bounded phase gives a much longer estimated duration (164–2009 years; median 681 years; Figure S2). This is not an artefact of smaller sample size: all possible combinations of three dates drawn from the seven pig samples produce much shorter duration estimates when modelled in the same way (Figure S2; the highest median duration for any set is 117 years).
Stratigraphic modelling on the assumption that the pig remains from above the YCB were not redeposited has good overall agreement (Amodel: 82.2; Figure S3: Model 3), suggesting this interpretation of the stratigraphy is plausible. This model increased the median estimate for the duration of pig deposition to 126 years (0–247 years; Figure S5: Model 3) with start and end boundaries of 2630–2470 cal BC and 2490–2350 cal BC respectively (Figure S3). Under this model, the construction date of the YCB is estimated at 2550–2470 cal BC (Figure S3).
If, by contrast, we assume that all samples from above the YCB may come from reworked deposits (Model 4, Figure S4), a likely scenario based on reconsiderations of the site’s stratigraphy (see below; for reviews, see Carlin Reference Carlin2017; Reference Carlin2018), we can only provide a terminus post quem for the construction of the YCB, estimated at 2560–2450 cal BC (Figure S5).
Zooarchaeology
Reviewing Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1986) fusion analysis to include all supplied data and using a standardised methodology for interpreting this (Tables S7–9), confirms her interpretation that the peak of slaughter is between 2 and 2.5 years, although this drops from 57 % to 52 %, and there is a greater proportion of older individuals present, with 22 % versus 13 % slaughtered at an age of over 3.5 years. This also confirms her overall assertion that based on fusion the majority of pigs were killed in the period May–November. Closely examining the data for distal tibiae, metapodia, and calcanei permits further refinement. For the tibia, which is the first to fuse at 24 months, only 7.8 % (11 out of 142) were unfused. For the metapodials, fusing up to three months later at 24–27 months, 17.7 % (3 out of 17) were unfused while for the calcaneus, fusing at 24–30 months, 69.6 % (48 out of 69) were unfused. This suggests that the majority of individuals in the 2–2.5-year age group were culled between 27 and 30 months, which, assuming a May farrowing, indicates a peak of slaughter between August and November.
In Van Wijngaarden-Bakker’s analysis of third molars she added two or three months to the estimated ages proposed by Higham (Reference Higham1967), resulting in 35 % of slaughter in October to December and 33 % in December to April. This is at odds with her fusion data, which showed a peak in May–November. There was also some ambiguity around the recording of third molars with wear beyond Higham’s stage 23, and no toothwear data for earlier slaughter ages. Since Van Wijngaarden-Bakker’s analysis, Grant’s (Reference Grant1982) work has refined the recording of Higham’s stages and these, and the associated age groups, have been routinely used. By applying them here and removing the additional growth period added by Van Wijngaarden-Bakker, the seasonality data suggests 68 % of slaughter in the period August to February rather than October to April.
While fusion data are generally considered to have less resolution than tooth data, these methods can be combined to support each other. The revised toothwear data indicate a peak in culling from August to February, while the revised fusion data indicate August to November. Together these suggest a peak August to November slaughter period for pigs, fitting the expected slaughter pattern of September to January (Parsons Reference Parsons1962). There is also evidence in Van Wijngaarden-Bakker’s data (Reference Van Wijngaarden-Bakker1986) for pigs at Newgrange being slaughtered at other times of the year, including in the later winter/early spring and during the summer, indicating that the site was being used throughout the year. Nevertheless, these data can be interpreted in conjunction with the isotopic data to further constrain and nuance the seasonality of the site (see below).
Discussion
Chronology
Radiocarbon dates from seven pig (this study) and three cattle (Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a) bones provide a clear answer to RQ1, that the fauna excavated by O’Kelly was not contemporaneous with the use of Beaker pottery in Ireland (c. 2450–2050 BC). When all 10 dates are considered, we see activity over a two-century period sometime between 2760 and 2480 cal BC (Figure 6 and 7: Model 1). These samples represent material excavated in both the eastern and western units, indicating that this temporal window captures activity across the site. When the pig dates are treated as a group, these indicate a short phase of depositional activity during the late 27th century to early 25th century cal BC (Figure 7: Model 2). Contra O’Kelly et al.’s (Reference O’Kelly, Cleary, Lehane and O’Kelly1983) hypothesis suggesting these faunal materials reflect ‘Beaker-period occupation’, this shows that these bones pre-date the adoption of this ceramic type and were contemporary with the use of Grooved Ware at Newgrange (Carlin Reference Carlin2017; Reference Carlin2018). We acknowledge that our sample size remains small, and that future dating evidence could show greater variation.
In answer to RQ2, the cohesiveness of these tightly clustered radiocarbon dates does not provide further evidence for stratigraphic mixing. Instead, they represent a unified chronological horizon pre-dating 2450 BC which may seem at odds with the multi-period nature of other deposits outside the tomb, including artefacts such as Beaker pottery that post-date 2450 BC. However, it is our understanding that the specimens from under the YCB were found in their primary context, having been protected from later disturbance by this bank, while those from over the bank seem to have been subjected to redeposition. This is based on Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1986) observation of concentrations of anatomically adjacent elements, especially vertebrae and scapulae, suggesting the deposition of intact portions of pig skeletons under the YCB. She noted that the pigs above the bank showed broadly similar but less pronounced patterns of concentrations of these elements. In contrast, those from the multi-period deposits where the bank was absent were far more disconnected. While the excavation records do not enable us to add further information to this, our Model 2 results suggest that pigs excavated from deposits both above and below the YCB were slaughtered over a very short, potentially contemporaneous, window of time and that those above were subject to later disturbance. In this scenario, the estimated construction date range for the YCB based on pig dates in Model 3 (Figure S3), which assumes that deposits from below and above the YCB were both in their primary contexts, may dramatically over-constrain the timeframe for construction of the YCB. Larger-scale dating of the Newgrange faunal assemblage has potential to add nuance to this narrative.
By contrast, our radiocarbon results are well suited for defining a terminus post quem for the YCB’s construction. Model 4, which allows for the possibility that pig samples from above the YCB are in secondary contexts — while still assuming that their initial deposition was part of the same episode of activity as the other pig samples — indicates that the YCB’s construction began after 2560–2450 cal BC (95 % probability). This terminus post quem is within the period when Grooved Ware was still in use and is before the estimated date for the adoption of Beaker pottery. This is at odds with the presence under the YCB of so-called ‘rusticated ware’, ‘Neolithic-derived bowl forms’, and ‘Beaker-associated coarse ware’, which were all originally considered part of a contemporaneous Beaker assemblage (Cleary Reference Cleary1983). However, these may well represent earlier styles of pottery, given that other Newgrange ceramics which were also treated as Beakers have subsequently been recognised as Grooved Ware (Roche Reference Roche1995; Brindley Reference Brindley1999). That accords with the absence of radiocarbon-dated pigs that overlap with the use of Beakers, either below or above the YCB. However, this can only be resolved through future work, including further reassessment of the pottery. Below, we return to considering the implications of our results for understanding the construction and function of this bank.
Interesting patterns emerge when dates from pigs are considered separately from cattle. The very short event duration modelled for pigs compared to cattle has implications for species-specific activities, which addresses RQ3 regarding taxonomic or spatial patterns in human–animal interactions (Figures 6 and 7: Model 2). The pigs date to 2600–2440 cal BC with a median duration estimate of 25 years under the simplest model (Model 2) and, at an extreme, the determinations could plausibly reflect a single event (ie less than one year). That these pig remains come from a range of locations suggests that the event or series of events that created the pig-rich deposits at Newgrange was highly constrained in time (Figure 7: Model 2). With respect to documenting human–pig interactions at Newgrange, a key aim of this paper, we prefer Model 2 over the other models because it focuses specifically on the pigs and relies on no other assumptions.
It is not clear why cattle (n = 3), with a modelled median duration estimate of 678 years, should show more temporal spread than pigs (n = 7, median duration estimate of 25 years). While the larger sample size for pigs might have been expected to produce somewhat tighter duration estimates, even artificially reducing sample size to only three specimens to match the cattle (by sampling all possible combinations of three pig dates) still resulted in a maximum median duration estimate of only 117 years (Figure S2). We believe the difference between cattle and pigs reflects a real pattern in the temporality of human–animal interactions at the site (see Appendix S8) but note that additional dates from cattle are needed to explore this interspecific pattern further.
In contrast to the cattle, the floruit of pig dates suggests an intense phase of pig-related activities representing feasting. If we assume that the seven pig dates are representative of the pig-centric nature of the assemblage, given the MNI evidence that at least 200 pigs (>150 from the western unit alone) were slaughtered and deposited in the ‘Beaker layers’, the shortest extreme of the modelled durations (ie an event duration of one year) seems less plausible. According to Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1986) calculations, based on MNI, these pigs represent a minimum of about 12,000 kg of dressed meat and fat, and such an outlay of livestock being expended for a single event seems unlikely, although not impossible, particularly if not all the meat was eaten, and some was instead deposited, perhaps as offerings. Assuming feasting consumption of a generous 2 kg of meat per person per day this would equate to 6000 portions, or enough to feed over 850 people for a week (Appendix S9). It is nevertheless easier to imagine a scenario in which these pigs reflect episodic feasting events, involving the consumption of very considerable quantities of meat, which took place over a period of a few years to a few decades. Notably, the shorter duration (be it one, five, or 50 years) we have observed in these modelled dates complements other patterns in the distribution and isotope compositions of pig bones at the site. Dense concentrations of pig remains, including numerous associations of specific elements consistent with the deposition of articulated portions, as well as a specialised and yet broadly consistent set of husbandry practices (see below), suggest that at least some of these deposits derive from a relatively short time span.
The temporally constrained nature of the pig dates suggests that the feasting events or processes that led to the pig-rich deposits may have been connected to the subsequent construction of the YCB, and potentially both may have served related goals. They were preceded by the construction of the large timber enclosure, but they all form part of a dynamic series of intense episodes of Grooved Ware-associated activity at the front of the large passage tomb in the two centuries c. 2700–2450 cal BC, immediately following the closure of its entrance (Carlin Reference Carlin2017; Smyth et al. Reference Smyth, Geber, Carlin, O’Sullivan, Griffiths, Hofmann, Cummings, Bjørnevad-Ahlqvist and Iversen2025). Evidence for intense midwinter pig feasting events like those at Newgrange has been identified at various timber or earthen monuments, including Durrington Walls, Mount Pleasant and West Kennet palisaded enclosures in southern Britain (eg Craig et al. Reference Craig and Parker Pearson2015; Bayliss et al. Reference Bayliss, Cartwright, Cook, Griffiths, Madgwick, Marshall, Reimer, Bickle, Cummings and Hofmann2017; Greaney et al. Reference Greaney, Hazell, Barclay, Ramsey, Dunbar, Hajdas, Reimer, Pollard, Sharples and Marshall2020). These large-scale seasonal gatherings of people and animals also occurred for short durations during the 26th and 25th centuries and were directly associated with monument building activities and the use of Grooved Ware. The Newgrange hearths may well have played a role in the cooking of pigs during Grooved Ware-associated feasting activities, similar to examples within small buildings at Durrington Walls, where large-scale feasting remains were enclosed by a henge in the 25th century BC (Craig et al. Reference Craig and Parker Pearson2015).
The resemblance of the Newgrange YCB to the banks of nearby embanked enclosures or henges (their counterparts in Britain) suggests that the YCB was built to further encircle the exterior of the passage tomb; other passage tombs at Brú na Bóinne are enclosed in the same way (Cooney Reference Cooney2006; Carlin Reference Carlin2017; Davis & Rassmann Reference Davis and Rassmann2021). While the original extent of the YCB is hard to reconstruct, by circumscribing the tomb’s southern and western perimeter, this bank directly continues a longer sequence of incomplete enclosing actions around the monument. However, it remains unclear if the absence of the YCB from the excavated area east of the tomb’s entrance is because it was subsequently removed or simply never constructed. O’Kelly et al. (Reference O’Kelly, Cleary, Lehane and O’Kelly1983) suggested it may have been created specially to cover earlier features, such as the underlying stone hearths. More recently, these hearths have been regarded as evidence for Grooved Ware-associated activities because of their resemblance to those found within Orcadian dwellings (Richards Reference Richards2005; Smyth Reference Smyth2011). Both interpretations of the YCB can be combined, based on comparison with the contemporaneous Grooved Ware-associated practices within henges in Britain, which were built through communal acts of labour that enclosed and memorialised the location of recent past activities involving feasting and social gatherings (eg Brophy & Noble Reference Brophy and Noble2012; Pollard Reference Pollard2012).
Husbandry
Isotopic data reveal the extent to which Newgrange pig husbandry was focused on woodland resources (RQ4) and, in turn, offer insights on seasonality of deposition at the site (RQ5) and more broadly, animal husbandry across Ireland (RQ6). Because we found no significant differences between contexts for both isotopic compositions, all pig data are treated as one interpretive unit. We consider Newgrange pig isotopic compositions in two ways. First, we compare pig data with published cattle isotopic compositions from Newgrange (Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a). Cattle are herbivorous but are likely to have fed more on grass and foliage than tree mast, which is poisonous to them (Cedervall et al. Reference Cedervall, Johansson and Jonsson1973; Dixon et al. Reference Dixon, McPherson, Rowland and Maclennan1979; González-Barrio et al. Reference González-Barrio, Truchado, García-Villalba, Hervás, Frutos, Espín and Tomás-Barberán2012). This means that cattle can provide a baseline against which to compare more ecologically and dietarily flexible pigs (see also Appendix S3). Second, we consider the Newgrange pig data within the wider corpus of isotope evidence for pig husbandry spanning the Neolithic to post-medieval periods in Ireland (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025).
The relationship between Newgrange pig and cattle δ 15N, with pigs producing a mean δ 15N value that is 0.7 ‰ lower than that of cattle, may seem surprising. This is because pigs and cattle are omnivores and herbivores, respectively, and δ 15N has traditionally been seen as an indicator of trophic position, such that animals incorporating meat into their diets are expected to show higher δ 15N than animals that do not (DeNiro & Epstein Reference DeNiro and Epstein1981). This serves to underscore that animal protein, including from aquatic sources, was likely unimportant in the diets of pigs slaughtered at Newgrange. The question remains, however, of why pigs, even those that were largely herbivorous, should have lower average δ 15N values than cattle at the same site. Acknowledging the many complexities of nitrogen sources and cycling (Szpak Reference Szpak2014), we consider three possible explanations in Appendix S10. We find that while it is not possible to establish definitively whether these animals were raised in the same area(s), and therefore the extent to which their δ 15N values should be comparable, it is parsimonious to proceed on basis that they were.
It is worth reviewing key temporal and biological dimensions of our interpretive context. Given the timing of mast abundance in Ireland’s woodland habitats (masting occurring in autumn, September–October; Kelly Reference Kelly1997) and the refined slaughter profile for pigs at Newgrange described above (potentially starting as early as August, but with a peak likely focused later in the year and completed by the end of the calendar year) we would expect isotope compositions of mast-fed pigs to show 13C enrichment especially in those slaughtered shortly after a period of mast-driven weight gain. There are two reasons for this. First, because mast as a non-photosynthetic tissue is 13C enriched, relative to foliage from the same plants (Cernusak et al. Reference Cernusak, Tcherkez, Keitel, Cornwell, Santiago, Knohl, Barbour, Williams, Reich and Ellsworth2009), there should be a clear signal in the bone collagen of mast consumers (Guiry et al. Reference Guiry, Orchard, Royle, Cheung and Yang2020; Reference Guiry, Orchard, Needs-Howarth and Szpak2021), with a difference of +2 to +4 ‰ being common (eg Sarà & Sarà Reference Sarà and Sarà2007; Guiry et al. Reference Guiry, Orchard, Needs-Howarth and Szpak2021).
Second, while the isotopic compositions of bone collagen are generally thought to reflect long-term (multiyear, or multidecadal) averages of consumers’ diets, these signals will be weighted toward periods of faster growth (Hobson & Clark Reference Hobson and Clark1992; Hyland et al. Reference Hyland, Scott, Routledge and Szpak2021). This will particularly be the case for animals slaughtered before or soon after growth plateaus (Warinner & Tuross Reference Warinner and Tuross2010). Pigs at Newgrange were largely dispatched between 27 and 30 months and, compared to older skeletally mature pigs, will have isotopic compositions that are more influenced by recently consumed foods. Food surfeits during the mast season are known to drive periods of accelerated weight gain (potentially more than doubling pig weights in a two- to three-month pannage period, if modern traditional breed Iberian figures can be transposed; eg Parsons Reference Parsons1962). This would be associated both with continued skeletal growth and considerable skeletal remodelling as the musculoskeletal system responds to changes in load bearing. Because the majority of pigs at Newgrange appear to have been slaughtered during the autumn–winter, we would expect isotopic signals associated with mast feeding, which are being recorded during a period of accelerated skeletal growth and adjustment, to have been amplified relative to periods of dietary intake earlier in life when growth was slower. It is important to note that pigs that were dispatched later, in the following year, would be expected to have lower δ 13C values, reflecting proportionately more materials from subsequent (non-mast) dietary intake.
Cattle from all time periods at Newgrange (n = 38) have δ 13C values consistently falling towards the lower end of the wider pan-Irish cattle isotope dataset (n = 388; Figure 4; Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a). At the other end sit cattle from early medieval sites (n = 71), when cattle husbandry had become a key driver of land management practices (Kelly Reference Kelly1997; Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a). Because cattle grazing and foddering focuses on foliage, and does not preferentially incorporate mast (Cedervall et al. Reference Cedervall, Johansson and Jonsson1973; Dixon et al. Reference Dixon, McPherson, Rowland and Maclennan1979; González-Barrio et al. Reference González-Barrio, Truchado, García-Villalba, Hervás, Frutos, Espín and Tomás-Barberán2012), the primary driver of δ 13C variation in Ireland is likely to be the canopy cover (Bonafini et al. Reference Bonafini, Pellegrini, Ditchfield and Pollard2013; for review see Guiry et al. Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a). In that context, lower cattle δ 13C at Newgrange suggests that many of these individuals were husbanded in (or with resources from) environments with denser canopy cover, likely woodland habitats, though natural landscape variation must also be expected. By contrast, cattle from the early medieval period, which represent animals more often grazed on open lands, provide a reference point for the highest δ 13C values that would be expected from non-mast feeding herbivores.
These patterns offer two important anchor points for interpreting pig data. First these data confirm that woodland habitat was likely being used for the husbandry of animals that were slaughtered at Newgrange. Second, cattle provide a baseline for the range of δ 13C variation expected in domestic herbivorous diets that do not incorporate tree mast. Using this interpretive framework, we can now address RQ4, which asks whether pig husbandry at Newgrange was, as hypothesised by McCormick (Reference McCormick, Murphy and Whitehouse2007), strongly woodland oriented. Our finding that pigs have significantly higher δ 13C relative to cattle at Newgrange, and even compared to cattle from the early medieval period (Figure 4) supports this, pointing to a husbandry regime in which pigs consumed large amounts of tree mast before slaughter (Appendix S11).
Fungus consumption has also been considered in previous research (Hamilton et al. Reference Hamilton, Hedges and Robinson2009; see Appendix S3 for further discussion). Pigs foraging in woodland habitats often have access to fungi, and these could be 13C enriched relative other C3 foods. However, fungi are also typically 15N enriched relative to local plants with which they have symbiotic relationships (Hobbie & Högberg Reference Hobbie and Högberg2012; Szpak Reference Szpak2014; O’Regan et al. Reference O’Regan, Lamb and Wilkinson2016). Therefore, given that our pig δ 15N values are on average lower than those of herbivorous cattle (Appendix S10), this explanation is unlikely to be a primary driver of the higher pig δ 13C values at Newgrange.
We can also examine how Newgrange pig isotopic compositions compare with a wider dataset across the Irish Holocene (Figure 5B; Knudson et al. Reference Knudson, O’Donnabhain, Carver, Cleland and Price2012; Guiry et al. Reference Guiry, Beglane, Szpak, Schulting, McCormick and Richards2018; Reference Guiry, Beglane, Szpak, McCormick, Teeter, Cheung and Richards2023a; Reference Guiry, Beglane, McCormick, Tourigny and Richards2025; Madgwick et al. Reference Madgwick, Grimes, Lamb, Nederbragt, Evans and McCormick2019a; McKenzie et al. Reference McKenzie, Murphy, Guiry, Donnelly and Beglane2020). Isotopic compositions from Neolithic and Early Bronze Age pigs are extremely rare, but the small pool of existing data shows wide ranging δ 13C and very low δ 15N, suggesting diverse husbandry practices, including both woodland and non-woodland oriented husbandry, with no evidence for diets with high quantities of animal protein (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). Within this context, pigs from Newgrange show less variable diets (Figure 5C) with a similar degree of herbivory (Figure 5D) and a greater emphasis (ie slightly higher mean δ 13C) on mast feeding (Figure 5A). The mast-focused diets of Newgrange pigs are particularly distinctive when compared with pigs from the Middle–Late Bronze and Iron Ages (n = 119; Figure 5A), which show more variable diets (Figure 5C), with a trend to lower δ 13C and a general uptick in δ 15N. This increase in δ 15N has previously been observed in published Irish pig isotopic compositions and is thought to be at least partly explained by broadscale human impacts on the cycling of nitrogen in soil brought on by changing land management practices in this period (Guiry et al. Reference Guiry, Beglane, Szpak, Schulting, McCormick and Richards2018; for an English example see Madgwick et al. Reference Madgwick, Mulville and Stevens2012). In that context, as with Newgrange and other earlier prehistoric sites, pigs from Middle–Late Bronze and Iron Age sites were largely herbivorous (Figure 5D). Pigs from the medieval and post-medieval periods show more extreme variation (Figure 5C) centered on moderate δ 13C but higher δ 15N values (Figures 5A and 5D). The extent to which δ 15N is elevated for this latter group reflects prevailing increases in animal protein consumption (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025).
Against this backdrop, we can now address RQ6, which queries whether there are parallels between the zooarchaeological evidence for an unusual abundance of pigs at Newgrange and isotopic evidence for specialisation in how those pigs were raised. We acknowledge that the extent to which we can address this is limited by generally smaller sample sizes at comparison sites, although broad insights can still be gained by comparing the Newgrange results with pig populations at wider spatial and temporal scales. For contemporaneous and later sites, isotopic compositions of Newgrange pigs show on average higher (statistically significant for all but the Neolithic/Early Bronze Age group) δ 13C values (Figure 5A). It is therefore clear that at Newgrange a unique pattern in pig husbandry was practised, involving a stronger focus on woodland resources than was common at other times or in most areas across Ireland for which we have data. Moreover, the supplemental animal protein sources (eg food scraps, agricultural byproducts) which appear increasingly important to Irish pig diets through the historical period (Figure 5D) are notably absent at Newgrange, as has been observed more widely for Neolithic/Early Bronze Age pigs across Ireland (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025).
Pannage and woodlands
These comparisons between pig diets at Newgrange and broader spatiotemporal trends serve to further underscore the significance of woodland resources. In his review of early medieval texts on Irish farming, Kelly (Reference Kelly1997) found that pigs maintained a high profile across major textual sources. These highlight the antiquity of associations between pigs and oak woodlands in Irish agriculture (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). Kelly’s (Reference Kelly1997) analyses of law texts, reflecting ecological knowledge on pig husbandry, indicate that, while pigs could be fed a variety of foodstuffs, mast was widely thought to be the most important. An excellent example is seen in laws that establish a legal distinction between acorn-fed pigs (muca for mesruth) and those raised on any other food (muca dema) (Kelly Reference Kelly1997). By the early medieval period, pig husbandry practices were so deeply entwined with oak woodland in Irish thinking that they had become proverbial. A notable example from the tale Togail Bruidne Da Derga (the destruction of Da Derga’s hostel), highlighted by Kelly (Reference Kelly1997), is the phrase ‘a pig which dies before the acorn crop’ (‘mucc remi-thuit mess’) which is used as a metaphor for a lost opportunity.
This historical context helps to further constrain isotopic interpretations. Given the entrenched connections between pigs and oak woodland in the early medieval period and given that the natural habitat of wild boar is woodland, it is possible that these concepts represent older traditional ecological knowledge. Combining documentary and isotopic data from the Iron Age and medieval periods (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025), it is clear that pig δ 13C values from these periods represent animals that, on average, had diets partly, or largely, focused on mast.
That Newgrange pig δ 13C values are on average higher than those from these later time periods is telling (Figure 5A; Appendix S12; Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025) and suggests that pig husbandry at Newgrange was even more closely tied to woodland resources than in the early medieval period, when the pig–pannage connection was first documented in Ireland. However, given the consistency in the revised fusion and tooth age-at-death estimates, higher δ 13C values could simply reflect highly regularised seasonal slaughter shortly after pannage-driven weight gain. Even so, it is tempting to suggest that the traditional ecological knowledge codified in the early medieval period is rooted in practices from a much deeper past. Indeed, based on the isotopic patterns observed in Newgrange pigs, it seems likely that the meaning of the early medieval proverb highlighted by Kelly would be easily recognised by those feasting at Newgrange 3500 years earlier.
Given that both the cattle and pigs slaughtered at Newgrange in the earlier 3rd millennium BC were husbanded in very specific ways that were intimately connected with woodlands, it is relevant to consider where these woodland habitats were located. Related to this, significant oak resources would have been required for the ongoing construction of timber monuments within Brú na Bóinne between c. 3000–2450 BC (Davis & Rassman Reference Davis and Rassmann2021). One might presume that pigs at Newgrange were being fed in dense oak woodland nearby. While current paleoenvironmental data lack sufficient detail to conclusively address this (Appendix S2), there is no indication that suitable mast-rich woodlands were present in the immediate area. Instead, Newgrange was seemingly in a largely open landscape with a pastoral character, generally regarded as prime land for mixed farming (Cooney Reference Cooney2000; Davis et al. Reference Davis, Lewis, Turner, Foster, Ostericher, O’Donnell, Collins, McClatchie, Allen and McCormick2017).
This suggests that the woodland-oriented husbandry practices we have observed reflect at least some degree of animal movement, a possibility that was also suggested by Van Wijngaarden-Bakker (Reference Van Wijngaarden-Bakker1986) for cattle. As detailed above, both the cattle and pigs at Newgrange display very specific age structures. While this may be taphonomic, it could also indicate that the animals consumed at Newgrange were neither born nor raised there. If they did come from a wider catchment, then the cohesive approach to their husbandry is even more striking. Such a possibility is supported by the isotopic evidence from the pigs slaughtered during midwinter feasting at Durrington Walls, which show high δ 13C values like those observed at Newgrange, but which also have other isotopic compositions that suggest diverse geological origins (Madgwick et al. Reference Madgwick, Lamb, Sloane, Nederbragt, Albarella, Parker Pearson and Evans2019b). There, the interpretation was that pigs were brought to Durrington Walls from up to 100 km away by various communities who assembled for midwinter feasting and monument building activities. It was also suggested that the number of pigs being raised specifically for midwinter feasting may have been so large that they could not be supported by locally available resources and were instead driven from settlements to woodlands specifically to be fattened on mast, before being brought to Durrington Walls.
There is considerable overlap in the evidence for animal husbandry and mobility practices, as well as short-lived feasting activity and associated monument building observed at Durrington Walls and Newgrange during the 26th and 25th centuries. However, this forms part of a much longer-term pattern of assembly of people, animals, and materials at Newgrange and Brú na Bóinne, at certain times of the year. At least five centuries previously, the Brú na Bóinne passage tombs incorporated multiple types of local and non-local stone and other materials brought from various places up to 40 km away by those who seasonally journeyed to participate in ongoing construction and ceremonial activities (Cooney Reference Cooney2000; Brady 2007; Reference Brady2018; Eogan & Cleary Reference Eogan and Cleary2017; Carlin et al. Reference Carlin, Smyth, Frieman, Hofmann, Bickle, Cleary, Greaney and Pope2025). As discussed below, Newgrange and Knowth continued to be a focus for large-scale gatherings and ceremonial activity until at least c. 2450 BC. During this time, these monuments and their surroundings saw an ongoing series of changes: tomb construction ceased by 2900 BC, whereas earthen or timber monument building peaked after 2700 BC. Yet, many aspects of the passage tomb-associated ceremonial activity, including communal gatherings, persisted until at least c. 2450 BC, (Carlin Reference Carlin2017; Eogan & Cleary Reference Eogan and Cleary2017). It would not be surprising if pigs, cattle, and oak timbers were also brought there in significantly increased numbers during the first half of the 3rd millennium BC. This is an issue requiring further work that can be readily addressed through other, widely available isotopic techniques and additional radiocarbon dating (see below).
Winter solstice at Newgrange after 2700BC
The temporality of human–pig–environment relationships has implications for our understanding of broader cultural and temporal trends at Newgrange. We have not generated comparable isotopic data from fauna contemporaneous with the construction and use of the large tomb at Newgrange (c. 3305–2700 BC; Lynch et al. Reference Lynch, McCormick, Shee Twohig, McClatchie, Molloy, Schulting, OCarroll and Sternke2014) so it is not possible to evaluate whether woodland-focused pig husbandry represented a new development during the 25th or 26th centuries BC or, more likely, was a continuation of even older traditions (Guiry et al. Reference Guiry, Beglane, McCormick, Tourigny and Richards2025). However, by considering the cycle of pig husbandry, we can address RQ5 (seasonality of site use) to explore a core theme — the winter solstice — around which activity at the site had been organised.
Considering the ways in which these isotopic analyses build on and refine existing zooarchaeological narratives is key to informing this exploration. The importance of mast supports Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1986) assessment that most pigs were dispatched within a six-month window in the autumn–winter seasons. While her methods could not be more specific about when this occurred, our isotopic analyses and refinement of the toothwear and bone fusion data suggests that slaughter was shortly after mast feeding had propelled pigs to their peak annual weight. Taking the medieval pannage season (September–October; Kelly Reference Kelly1997) as a benchmark, this suggests that pigs would have been slaughtered around the turn from autumn to winter, likely in the November to December window. We acknowledge slaughtering pigs at this time of year may reflect secular and/or economic factors, such as cooler weather facilitating easier meat preservation and the need to cull animals before they become harder to feed in winter, but as noted above, the scale, spatial patterning, and tight temporality of the pig assemblage at Newgrange all strongly suggest this pattern reflects a broader cultural phenomenon.
If we accept, as most now do (Mount Reference Mount1994; McCormick Reference McCormick, Murphy and Whitehouse2007; Carlin Reference Carlin2018), that these pigs reflect ritual, ceremonial, or at least social, feasting, rather than settlement, then it is difficult to ignore the timing of these activities. Together these lines of evidence suggest that people were physically present for feasting events leading up to and incorporating the winter solstice between the 26th and 25th centuries BC. The extent to which this directly echoes aspects of Newgrange’s construction and use (Hensey Reference Hensey2015), especially the gathering of people for the winter solstice, seems more than a coincidence. It is also difficult to ignore the location of these feasting remains outside the entrance. This deposition seems to represent a continuation of a long-standing tradition of enhancing the front of developed passage tombs through activities that emphasised their entrances (eg Carlin Reference Carlin2017). It is, therefore, reasonable to suggest that in the context of Newgrange, the greater concentration of faunal remains near the entrance relates directly to the celebration of the winter solstice, a phenomenon that had been a defining feature of this monument’s design, construction, and use since 3300 BC. Even though the sealing of the tomb c. 2700 BC prevented the beams of the winter solstice sunrise from being witnessed as they illuminated the passage, the solstice seems to have remained important. In that context, and addressing RQ5, we offer an alternative hypothesis for the origins of this faunal assemblage: that these pig remains reflect winter solstice-oriented ‘ceremonial’ activities outside the Newgrange tomb for at least two centuries (2700–2450 BC) later than previously thought.
This new evidence for the persistence of midwinter gatherings at Newgrange into the 3rd millennium BC confirms the extent to which emerging cultural influences from across the Irish Sea, such as Grooved Ware-associated feasting were incorporated within the Irish passage tomb tradition (see Carlin Reference Carlin2017; Carlin & Cooney Reference Carlin, Cooney, Card, Edmonds and Mitchell2020). Our results demonstrate the evolving nature of activity at Newgrange throughout this millennium, indicating continuity of both place and practice; Newgrange certainly did not go into decline either with the closure of the tomb, or the adoption of Grooved Ware in Ireland. Instead, activities continued at Newgrange during much of the 3rd millennium BC in much the same ways as beforehand, often directly referencing the pre-existing passage tomb monuments (Carlin & Cooney Reference Carlin, Cooney, Card, Edmonds and Mitchell2020). This includes concentrations of ceramics found outside the entrance (O’Kelly et al. Reference O’Kelly, Cleary, Lehane and O’Kelly1983) and in the timber structures that enclosed another nearby tomb (Site Z), resulting from Grooved Ware-associated feasts during communal gatherings and ceremonial undertakings between 2700–2450 BC (for reviews, see Carlin 2017; Reference Carlin2018). There is a growing body of evidence, albeit much of it from geophysical and aerial survey, and therefore undated, suggesting that activity around Newgrange, including monument construction using oak timbers and/or earth rather than stone, escalated considerably between c. 3000–2450 BC. Furthermore, some of these structures and enclosures also display a concern with solstice alignments (Carlin & Cooney Reference Carlin, Cooney, Card, Edmonds and Mitchell2020; Davis & Rassmann Reference Davis and Rassmann2021). It is therefore reasonable to suggest that those who continued to occupy this landscape were aware of the significance of the winter solstice at Newgrange and maintained a concern for solar events.
If this is true, another question emerges — when did this knowledge disappear? This has been explored by many researchers (Carey Reference Carey1990; Stout & Stout Reference Stout and Stout2008; Waddell Reference Waddell2014; Hensey Reference Hensey, Papadopoulos and Moyes2017). O’Kelly (Reference O’Kelly1982) considered sun-oriented themes in medieval narratives such as De Gabáin int Síde (the taking of the otherworld mound) and Tochmarc Étaíne (the wooing of Étaín) as potential evidence for the preservation of knowledge of Newgrange’s solar orientation. In that context, he asked if it could be that the tomb builders also ‘planted the seeds of Irish oral literature’, even pondering whether such medieval narratives could now be seen ‘not as a window on the Iron Age but as one on the Late Neolithic?’ More recent scholarship has highlighted potential pitfalls in transposing knowledge from these medieval narratives, which may at least partly represent coeval inventions, into the prehistoric past (Gleeson Reference Gleeson2020). Acknowledging this issue, our findings still show that concentrated activities, including feasting, occurred around the time of the winter solstice, centuries after the monument was sealed off. It is striking to reflect that such activities continued some generations after the last people experienced the solstice phenomenon from within the monument, suggesting a considerable degree of collective memory.
Conclusion
The faunal assemblage at Newgrange is one of the largest to be associated with any Irish passage tomb and holds potential to contextualise the chronology and nature of human activities at the site (Carlin Reference Carlin2018). In the decades since Van Wijngaarden-Bakker’s (Reference Van Wijngaarden-Bakker1974; Reference Van Wijngaarden-Bakker1986) work, this assemblage has remained silent, in part due to issues with chronological uncertainty and methodological limitations of zooarchaeology. A systematic dating programme to further clarify the chronology of the fauna and the sequence of activity is much needed, and some of this work has already begun (Smyth et al. Reference Smyth, Geber, Carlin, O’Sullivan, Griffiths, Hofmann, Cummings, Bjørnevad-Ahlqvist and Iversen2025; Carlin et al. Reference Carlin, Griffiths, Kanne and Smythforthcoming). However, Van Wijngaarden-Bakker’s core findings remain: the assemblage shows that many pigs were slaughtered on site in the autumn–winter months.
It has been previously argued that these deposits reflect feasting events associated with the use of Beaker pottery (Mount Reference Mount1994; McCormick Reference McCormick, Murphy and Whitehouse2007) but we have shown that the faunal remains largely represent an earlier floruit of feasting activity between the 26th and 25th centuries BC. This reflects a continued concern with the annual solstice event, which was so central to the tomb’s construction and use. It also aligns closely with the wider body of evidence for a continuity of practice and traditions of communal gatherings and ceremonial activity both at Newgrange and other passage tombs until at least c. 2450 BC, but probably beyond this (Carlin Reference Carlin2017).
People feasting at Newgrange were enmeshed in networks with other people, places, animals, plants, and things that operated at a local scale, but also extended across the Irish Sea as far as Orkney and southern Britain (Carlin Reference Carlin2017; Copper et al. Reference Copper, Sheridan and Whittle2024; Greaney et al. Reference Greaney, Hajdas, Dee and Marshall2025). These interconnections were materialised by the use of Grooved Ware pottery and other distinctive material culture that originated in different regions of Ireland and Britain but became widely used across much of these islands in the earlier 3rd millennium BC. These objects formed part of a suite of contemporaneous cultural innovations that enabled shared ideas and practices to be enacted in distinct ways at various buildings and monuments, including circular timber and earthen enclosures (Carlin & Cooney Reference Carlin, Cooney, Card, Edmonds and Mitchell2020). Our research highlights the strong parallels between the intense concentrations of activity associated with feasting event(s) at various henges and enclosures in southern Britain around 2500 BC, including Durrington Walls, Mount Pleasant and West Kennet palisaded enclosures, and at Newgrange. While there are clear similarities and differences in the character of the monuments at these places, contemporaneous large-scale seasonal gatherings at midwinter for monument building and feasting occurred at each. At Newgrange and Durrington Walls, at least, we can now say that the feasting was focused on pigs that had been specially fattened on mast in advance. In combination, this suggests that these places may have been strongly interconnected through highly fluid and intersecting webs of contact at this time, thereby resulting in shared practices and worldviews.
Further isotopic and other work could open new windows onto past human–animal–environmental relationships at Newgrange, advancing the husbandry-oriented interpretations we have made and developing new evidentiary frameworks for animal and human mobility. For instance, serial sampling of tooth dentine, which offers a diachronic perspective of diet at the intra-individual level (Sealy et al. Reference Sealy, Morris, Armstrong, Markell and Schrire1993), could be applied to pig canines to allow more in-depth consideration of the duration, to a sub-month level, and the number of years of pannage feeding that pigs experienced (Frémondeau et al. Reference Frémondeau, Cucchi, Casabianca, Ughetto-Monfrin, Horard-Herbin and Balasse2012; Guiry et al. Reference Guiry, Hepburn and Richards2016). Other techniques including compound-specific isotopic work such as analyses of single amino acids (McMahon & Newsome Reference McMahon, Newsome, Hobson and Wassenaar2019) and metabolomics (eg identifying acorn-derived polyphenols; Alldritt et al. Reference Alldritt and McCullagh2019) could help to further resolve these husbandry practices.
Additional biogeochemical indicators, such as sulfur, strontium, and oxygen isotope compositions could provide a framework for assessing pig mobility. Such work has begun to explore long-distance movements of animals deposited at other monumental sites in Britain and Ireland (Madgwick et al. Reference Madgwick, Grimes, Lamb, Nederbragt, Evans and McCormick2019a; Reference Madgwick, Lamb, Sloane, Nederbragt, Albarella, Parker Pearson and Evans2019b; Barclay & Brophy Reference Barclay and Brophy2021; Müldner et al. Reference Müldner, Frémondeau, Evans, Jordan and Rippon2022). In that context, it is interesting to note that the pig δ 13C and δ 15N values from Newgrange show less dietary variation than at other sites, such as the Iron Age Navan Fort, where there was evidence for feasting (Madgwick et al. Reference Madgwick, Grimes, Lamb, Nederbragt, Evans and McCormick2019a) and where isotopic proxies suggest a wide catchment for pig origins; however, that does not imply that all the pigs were local to Newgrange.
It is also worth highlighting that our primary interpretive link, connecting mast feeding with distinctive isotopic patterns, is relatively novel in the context of European prehistory (Appendix S13). Human–woodland interactions are important across Europe and the globe (eg Roberts et al. Reference Roberts, Hunt, Arroyo-Kalin, Evans and Boivin2017; Whitlock et al. Reference Whitlock, Colombaroli, Conedera and Tinner2018). The intimate connection between pigs and woodland in many European cultures (Szabó Reference Szabó and Rotherham2013; Weallans Reference Wealleans2013; Margetts Reference Margetts2022) means that archaeological isotopic perspectives on pig husbandry should provide a window on the evolving nature of those relationships through time. Following an early florescence of interest in the potential for isotopic analyses to evidence human–pig–woodland relationships (eg Hamilton et al. Reference Hamilton, Hedges and Robinson2009; Hamilton & Hedges Reference Hamilton, Hedges, Whittle, Healy and Bayliss2011; Hamilton & Thomas Reference Hamilton and Thomas2012; Madgwick et al. Reference Madgwick, Mulville and Stevens2012; Millard et al. Reference Millard, Jiménez-Cano, Lebrasseur and Sakai2013), a continued lack of clarity about what isotopic signatures for mast feeding should look like has served to relegate such interpretations to minor notes in more recent studies (though see Frémondeau et al. Reference Frémondeau, Cucchi, Casabianca, Ughetto-Monfrin, Horard-Herbin and Balasse2012; Reference Frémondeau, De Cupere, Evin and Van Neer2017; Souron Reference Souron, Melletti and Meijaard2017). In that context, we hope that our analyses, highlighting a clear connection between mast feeding and δ 13C signals, will help to reinvigorate this line of enquiry by encouraging reinterpretation of existing datasets and generating new interest in future analysis.
Acknowledgements
For sampling permission and logistical help, we thank Nessa O’Connor, Eimear Ashe (National Museum of Ireland), and Thorsten Kahlert (previously IT Sligo). We are also grateful to Matt Seaver and Alex Guglielmi (National Museum of Ireland) for their assistance with the recovery of key contextual details relating to the Newgrange faunal assemblage. We thank Finbar McCormick for comments on an early draft. Funding for EG included Wenner Gren Foundation, the University of British Columbia, and the Social Science and Humanities Research Council of Canada.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/ppr.2025.10063










 Open access
Open access